Hintergrundinformation
Toxische Cyanobakterien
Cyanobakterien [1] sind ein natürlicher Teil der Lebensgemeinschaften in unseren Gewässern. Ein Teil der Cyanobakterien kommt schwebend im Wasserkörper vor (planktisch), andere Arten besiedeln Oberflächen wie den Gewässergrund oder Steine (benthisch). Bei hohen Nährstoffkonzentrationen können sich Cyanobakterien extrem stark vermehren.
Cyanobakterien können neben dem Chlorophyll sogenannte akkzessorische Photopigmente enthalten, so z.B. das rote Phycoerythrin und das blaue Phycocyanin. Durch diese Pigmente entsteht die typische Färbung der verschiedenen Cyanobakterien, wodurch sie bei einem Massenauftreten das Wasser in unterschiedlichen Farben verfärben. Blau wirken Cyanobakterienblüten allerdings meist nur dann, wenn die Zellen stark geschädigt sind und das gut wasserlösliche Phycocyanin heraustritt.
Viele Arten von Cyanobakterien können Toxine produzieren, die bei Menschen und Tieren zu Vergiftungen führen können. Vor allem die im Süßwasser häufig vorkommenden potentiell toxischen Cyanobakterien wie Microcystis, Planktothrix, Aphanizomenon und Anabaena bilden unter günstigen Bedingungen Massenentwicklungen (“Algenblüten”), die zeitweilig fast alle anderen Phytoplankton-Arten verdrängen. Für die Beurteilung, ob Cyanobakterien in Badegewässern oder in der Trinkwasserversorgung eine Gefährdung darstellen, ist es wichtig, das Auftreten möglicherweise kritischer Gattungen schnell und sicher erkennen zu können.
Bei der Bewertung kommt es häufig nicht auf die genaue Artbestimmung an; meist genügt die Zuordnung zu einer Gattung, was von Mitarbeiterinnen und Mitarbeitern mit Erfahrung im Mikroskopieren nach einer Schulungsphase gut geleistet werden kann.
Als ersten Einstieg finden Sie auf dieser Seite eine Übersicht der am häufigsten vorkommenden potentiell toxischen Cyanobakterien. Weitere Quellen für einen Einstieg in die Thematik finden Sie im Literaturverzeichnis, im Internet, z.B. Fotos oder taxonomische Informationen und Literaturquellen.
Mit Hilfe dieser Quellen – und auch unter Hinzuziehung externer limnologischer Kompetenz – können Sie die wichtigsten Cyanobakterien in Ihrem Gewässer bestimmen und eine Liste für deren Bestimmung in den Folgejahren erstellen.
Häufige toxinbildende Cyanobakterien
Die nachfolgenden Cyanobakterien bilden in Mitteleuropa häufig Massenentwicklungen und können Cyanotoxine produzieren.
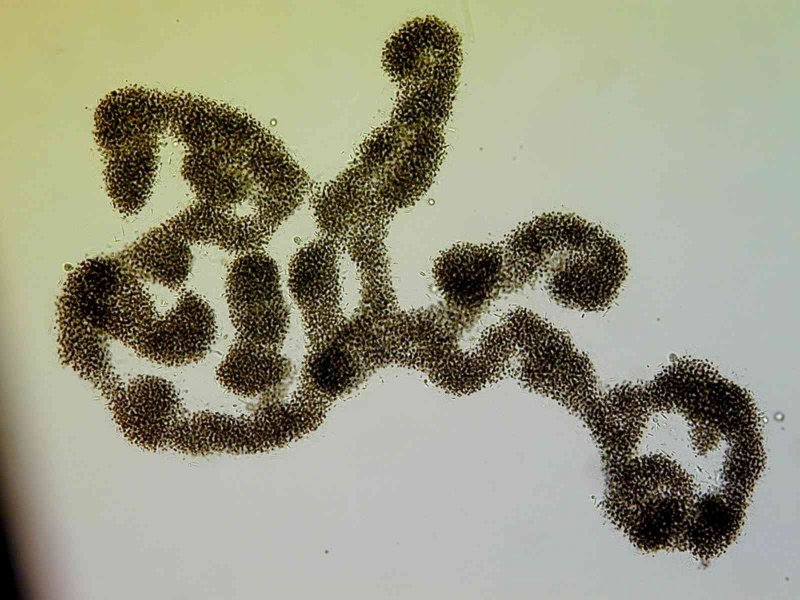
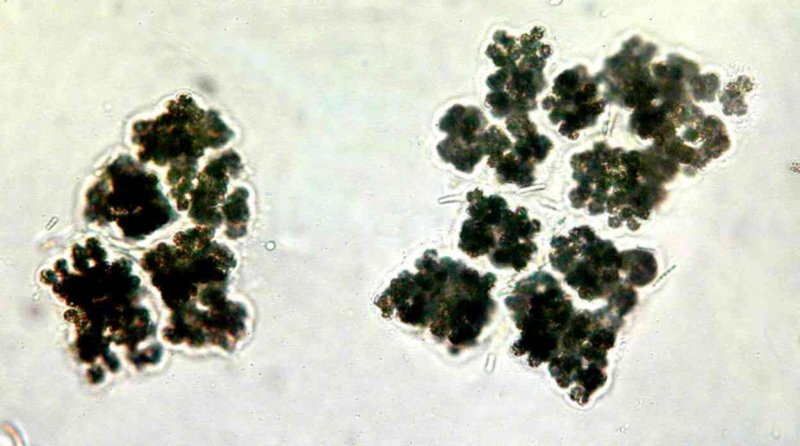
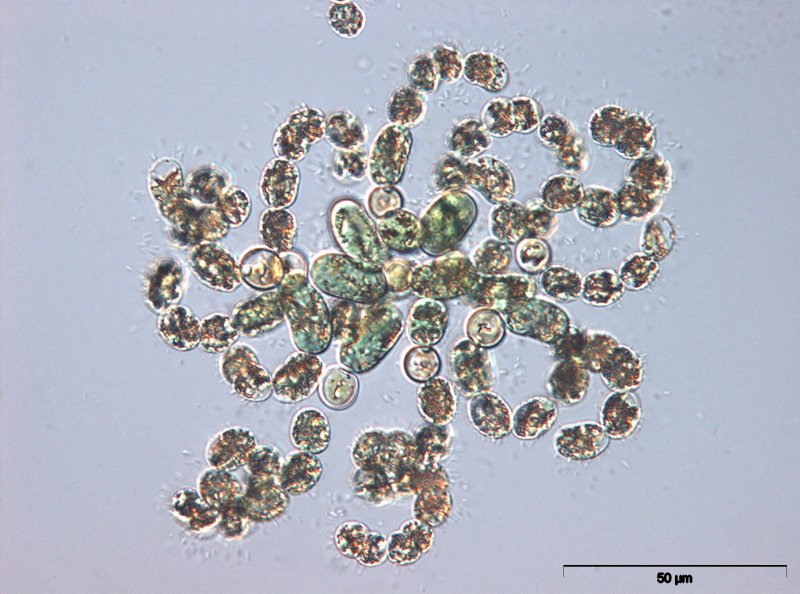
Microcystis
Die Gattung Microcystis ist weltweit im Süßwasser verbreitet. Ihre Einzelzellen sind kugelig bis ovoid mit einem Durchmesser von 2,5 – 8 µm und bilden große Kolonien, die oft mit dem bloßen Auge erkennbar sind. Durch Gasvesikel in den Zellen können die Kolonien aufrahmen und Schlieren oder gar dicke Schichten auf der Wasseroberfläche bilden. Die einzelnen Microcystis-Arten werden vor allem durch die Zellgröße sowie die Form der Kolonien unterschieden.
Die am häufigsten in eu- bis hypertrophen Gewässern auftretenden Microcystis_-Arten sind M. aeruginosa, _M. flos-aquae, M. viridis, M. ichthyoblabe und M. wesenbergii. Bei diesen Arten wurden häufig Microcystine nachgewiesen, andere der bekannten Cyanotoxine (z.B. Anatoxine, Cylindrospermopsine) dagegen bislang nicht gefunden. Bei hohen Nährstoffkonzentrationen kann Microcystis starke Massenentwicklungen bilden. In diesen Blüten können lokal sehr hohe Microcystinkonzentrationen vorkommen, die beim Verschlucken größerer Mengen einer solchen Zellsuspension akut gesundheitsgefährdend sind.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier
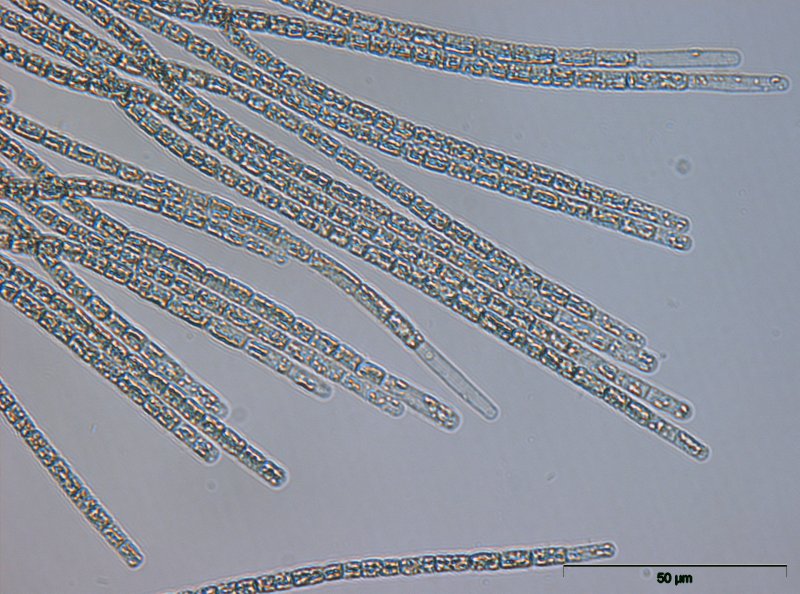
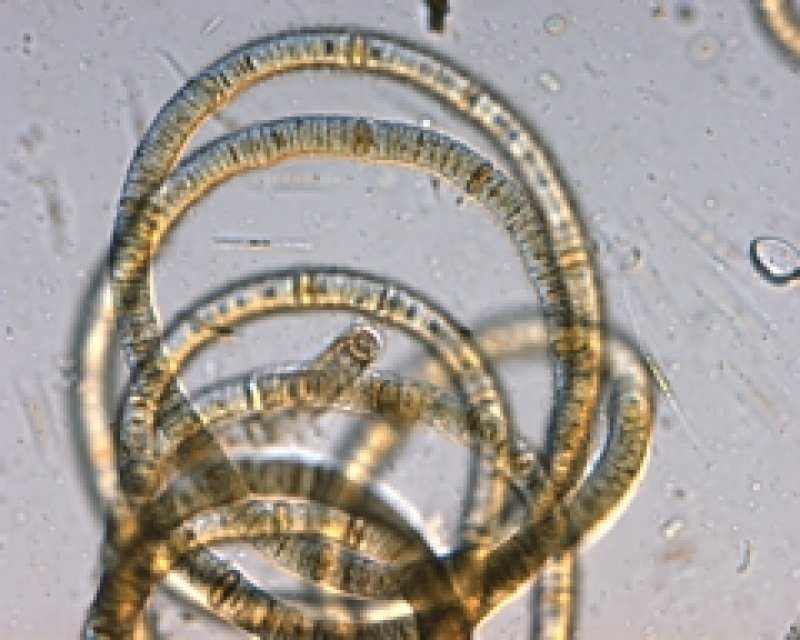
Planktothrix
Cyanobakterien der Gattung Planktothrix wachsen in langen Filamenten [2] mit Gasvesikeln. Ihre Zellen sind in der Regel breiter als lang mit Zelldurchmessern von 3,5 – 10 µm. Die Filamente sind in gewissem Maße mobil und können daher in der Trinkwasseraufbereitung durch Filter gelangen.
Microcystine sind die am häufigsten nachgewiesenen Cyanotoxine der Gattung Planktothrix, vereinzelt wurden auch Neurotoxine (Anatoxin-a und Saxitoxin) gefunden. Die bekanntesten Microcystinproduzenten dieser Gattung sind Planktothrix agardhii und Planktothrix rubescens, letztere wegen der roten Färbung auch “Burgunderblutalge” genannt. Diese beiden Arten sind weltweit verbreitet, kommen aber in unterschiedlichen Ökosystemen vor:
Planktothrix agardhii dominiert oft das Plankton in eu- bis hypertrophen flachen und deshalb häufig durchmischten Gewässern. Die Durchmischung bedingt, dass Planktothrix agardhii im Gegensatz zu anderen Cyanobakterienarten keine Schlieren an der Gewässeroberfläche bildet, sondern stets mehr oder weniger homogen im Wasser verteilt ist.
Planktothrix rubescens kommt dagegen in tiefen, thermisch geschichteten meso- bis eutrophen Gewässern vor, wo sie während der Sommermonate vor allem im Metalimnion [5] vorkommt, da ihre rote Pigmentierung eine optimale Lichtausbeute in dieser Tiefe bei gleichzeitiger Nutzung von nährstoffreichem Tiefenwasser erlaubt. Erst bei der Durchmischung der Gewässer im Herbst wird sie in andere Gewässerschichten verteilt, und dann kann es unter nährstoffreichen Bedingungen auch zu Oberflächenblüten kommen. Planktothrix rubescens kommt in manchen Gewässern ganzjährig vor, auch unter Eis.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier

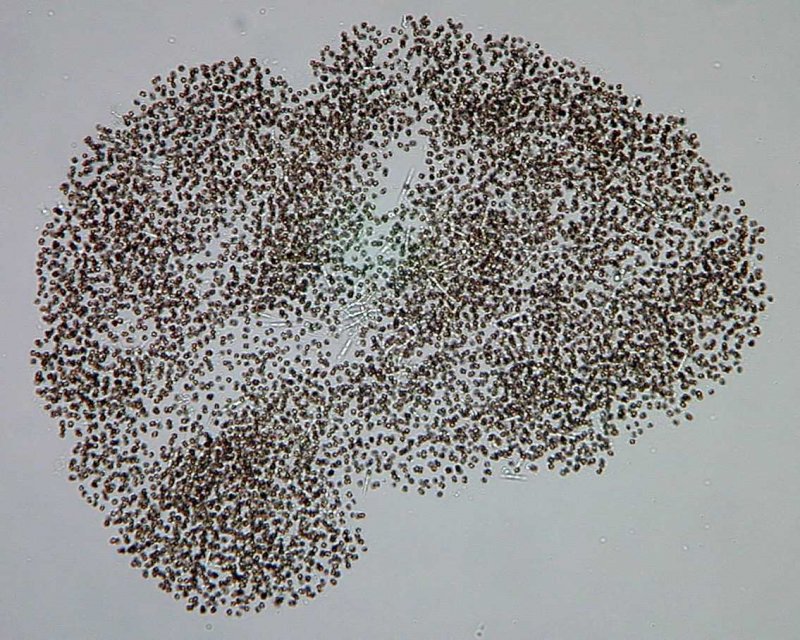
Aphanizomenon/Cuspidothrix
Vertreter der Gattung Aphanizomenon wachsen als Filamente (Zellfäden) mit 20 – 50 Zellen pro Filament [2] und einen Durchmesser zwischen 3 – 7 µm je nach Art. Zudem können in den Filamenten Heterocysten [3] und Akineten [4] vorkommen. Die einzelnen Filamente können sich bei Aphanizomenon flos-aqaue büschelweise zusammenlagern und so makroskopisch sichtbare Aggregate bilden, die im Wasser schweben und den Eindruck etwa von Lärchennadeln oder frisch geschnittenem Gras erzeugen und gelegentlich auch an der Gewässeroberfläche akkumulieren. Andere Arten kommen ausschließlich als Einzelfilamente vor, wie z.B. Aphanizomenon gracile oder Cuspidothrix (früher Aphanizomenon) issatschenkoi. Die meisten Vertreter der Gattung Aphanizomenon sind planktische Süßwasserbewohner, Aphanizomenon sp. kommt aber auch in der Ostsee vor.
Von den bekannten Cyanotoxinen wurden bislang bei Aphanizomenon Paralytic Shellfish Poisons (Saxitoxine), Anatoxin-a und Cylindrospermopsin, jedoch nie Microcystine nachgewiesen. Aphanizomenon gracile, Cuspidothrix (früher Aphanizomenon) issatschenkoi, Aphanizomenon flos-aquae und Aphanizomenon ovalisporum sind die am häufigsten mit Toxinproduktion in Verbindung gebrachten Arten.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten der Gattung Aphanizomenon finden Sie hier.
Anabaena (syn. Dolichospermum)
Die Arten der Gattung Anabaena bilden unverzweigte Filamente [2], in denen häufig Heterocysten [3] vorkommen, unter ungünstigen Bedingungen auch Akineten [4]. Die einzelnen Zellen (4-14 µm Durchmesser) sind kugelförmig bis zylindrisch. An den Zellgrenzen sind die Fäden stets eingeschnürt bis perlschnurartig. Die Filamente können gerade (z.B. Anabaena circinalis), gewunden (z.B. A. spiroides) oder zu Knäulen verwickelt sein (z.B. A. lemmermannii). Die Gattung Anabaena ist weltweit häufig im Süßwasserplankton verbreitet, jedoch gibt es auch Arten die im Brackwasser vorkommen und benthische Lebensformen (z.B. Matten auf Steinen).
Bei Vertretern der Gattung Anabaena wurden alle bekannten Cyanotoxine nachgewiesen: Microcystine, Cylindrospermopsin, Anatoxin-a, Anatoxin-a (s) sowie Paralytic Shellfish Poisons (Saxitoxine). Bei Massenentwicklungen kann es lokal durch Aufrahmungen an der Oberfläche zu hohen Toxinkonzentrationen kommen.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier
Nodularia
Die Vertreter der Gattung Nodularia kommen überwiegend in Brackwasser vor. Die toxische Art Nodularia spumigena bildet fädige Zellketten, die mehr oder weniger gerade, gebogen, oder auch unregelmäßig spiralig sein können. In den Filamenten [2] treten in regelmäßigem Abstand Heterocysten auf. N. spumigena tritt vorwiegend in Brackwasser und Salzwasser mit bis zu 30 % Salzgehalt auf und ist weltweit verbreitet. In der Ostsee bildet N. spumigena im Sommer regelmäßig Massenentwicklungen, wobei bei ruhigen Wetterlagen die Zellfäden aufgrund von Gasvesikeln auftreiben und ggf. an die Küsten getrieben werden.
N. spumigena kann das Lebergift Nodularin produzieren.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier


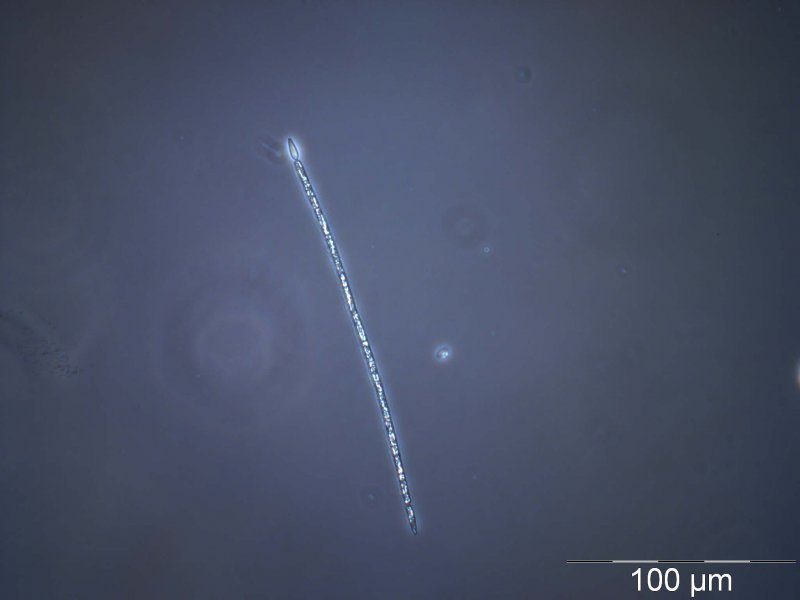
Cylindrospermopsis
Die Gattung Cylindrospermopsis tritt überwiegend in tropischen Gewässern auf. Auch Cylindrospermopsis raciborskii war ursprünglich in den (Sub)tropen verbreitet, allerdings hat sich diese Art in den letzten Jahrzehnten auch in gemäßigten Klimazonen ausgebreitet. C. raciborskii ist ein fadenförmiges Cyanobakterium meist mit einer charakteristischen lanzettförmigen Heterocyste [3] am Ende des Fadens. Unter schlechten Wachstumsbedingungen bildet diese Art Dauerzellen.
C. raciborskii kann Cylindrospermopsine und Paralytic Shellfish Poisons (Saxitoxine) produzieren.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier
Weitere toxinbildende Cyanobakterien
Auch bei den folgenden Gattungen wurde die Produktion von Cyanotoxinen nachgewiesen. Das Gefährdungspotential dieser Gattungen für Gewässernutzung und Trinkwassergewinnung wird aber als weniger relevant eingeschätzt.
Anabaenopsis
Die Gattung Anabaenopsis umfasst derzeit ca. 15 Arten filamentöser Cyanobakterien. Die Filamente [2] sind gerade, gebogen oder spiralig. In den Filamenten bilden sich Heterocysten [3] und Akineten [4]. Da die Filamente nach der Ausbildung von Hetereocysten häufig brechen, liegen die Heterocysten an den Enden der kurzen Filamente. Die Zellen sind zylindrisch.
In Vertretern der Gattung Anabaenopsis wurden vereinzelt Microcystine nachgewiesen.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier
Cylindrospermum
Die Vertreter der Gattung Cylindrospermum wachsen als Filamente [2], die feine oder kompakte Matten auf Oberflächen in wenig eutrophen Gewässern bilden. Die Filamente sind leicht gebogen bis stark gekrümmt. Die Zellen sind meistens zylindrisch. Heterocysten entwickeln sich nur an den Enden der Filamente, sind ovoid, oval oder konisch. Akineten [4] entwickeln sich nur in der Nähe der Heterocysten [3].
Bei manchen Arten der Gattung Cylindrospermum sind Anatoxine nachgewiesen worden.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier
Lyngbya
Die Vertreter der Gattung Lyngbya bilden dicke Filamente [2], die sich häufig zu festen Matten zusamenlagern. Die Filamente bestehen aus diskusförmigen Zellen, die immer breiter als lang sind. Heterocysten [3] und Akineten [4] fehlen. Lyngbya tritt weltweit auf, häufig im Brackwasser oder in marinen Lebensräumen.
Bei einigen Arten der Gattung Lyngbya wurden Lyngbyatoxine, Aplysiatoxine, Cylindrospermopsine und Paralytic Shellfish Poisons (Saxitoxine) nachgewiesen.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier
Nostoc
Die Gattung Nostoc umfasst Cyanobakterien, die kugelige oder hautartige Kolonien aus langen, unverzweigten Zellschnüren in einer gelatinösen Hülle bilden. Innerhalb der Zellketten kommen Heterocysten [3] und unter ungünstigen Wachstumsbedingungen auch Akineten [4] vor.
Bei einigen Arten der Gattung Nostoc, auch bei symbiontisch in Flechten lebenden Vertretern, wurden Microcystine oder Nodularine nachgewiesen.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier
Oscillatoria
Die Gattung Oscillatoria tritt häufig in Süßwasser, Brackwasser und marinen Lebensräumen auf. Es exisitieren planktische und benthische Vertreter. Oscisllatoria bildet lange unverzweigte Filamente [2]. Heterocysten [3] oder Akineten [4] treten nicht auf. Die Zellen sind immer breiter als lang und die Filamente wirken daher etwas wie Geldrollen.
Bei manchen Arten der Gattung Oscillatoria wurden Microcystine, Anatoxin-a, Homoanatoxin, und Aplysiatoxin nachgewiesen.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier.
Phormidium
Vertreter der Gattung Phormidium wachsen meist als Filamente [2] in Matten auf unterschiedlichem Substrat (Erde, Wasserpflanzen, nasse Steine etc.)
Phormidium-Arten können Anatoxin-a und Homoanatoxin-a produzieren, und waren oft die Ursache von neurotoxischen Vergiftungen von Tieren. Vereinzelt gibt es auch Hinweise auf Microcystinproduktion.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier
Raphidiopsis
Die Vertreter der Gattung Raphidiopsis wachsen als gerade, gewundene oder schraubenförmige Einzelfilamente [2], die am Ende spitz zulaufen. Sie bilden Akineten [4], aber keine Heterocysten [3]. Nur wenige planktisch lebende Süßwasser-Arten.
Als Toxinproduzenten sind Raphidiopsis mediterranea (Cylindrospermopsin, Deoxy-Cylindrospermopsin, Anatoxin-a, Homoanatoxin),Raphidiopsis curvata (Cylindrospermopsin, Deoxy-Cylindrospermopsin) und Raphidiopsis brookii (Saxitoxin) beschrieben.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier
Schizothrix
Die Vertreter der Gattung Schizothrix bilden Filamente [2], die meist von gelatinösen Scheiden umgeben sind, welche sich charakteristisch an den Enden verjüngen und abgestumpfte Spitzen bilden. Die Zellen sind zylindrisch und gewöhnlich länger als breit, am Fadenende sind sie überwiegend konisch. Heterocysten [3] und Akineten [4] fehlen.
Die Filamente bilden gewöhnlich Lager, z.B. auf Steinen, die auch in Größen vorkommen können, die mit bloßem Auge sichtbar sind. Diese Lager treten in vielen verschiedenen Erscheinungsformen auf, z.B. Lager mit Kalk inkrustiert, pelzig, etc.
Manche Arten der Gattung Schizothrix produzieren Aplysiatoxine.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier
Tychonema
Die Vertreter der Gattung Tychonema wachsen als gerade Filamente [2] mit einer Zellbreite von 5 – 16 µm. Die Filamente sind isodiametrisch mit abgerundeter Endzelle. Charakteristisch ist ihr durchscheinender Zellinhalt, ggf. erweiterte Thylakoide (“Vakuolen”) sowie die zentripetale Bildung der Querwände. Die beschriebenen Arten unterscheiden sich im wesentlichen durch die Zellbreite sowie den Lebensraum (planktisch, benthisch, tychoplanktisch, epiphytisch).
Die Bildung von Anatoxin-a ist für T. bornetii und T. bourrellyi nachgewiesen, für T. tenue wahrscheinlich.
Detaillierte Informationen zur taxonomischen Bestimmung der Arten dieser Gattung finden Sie hier